Canine Coat Colour Genetics& The Miniature Schnauzer
|
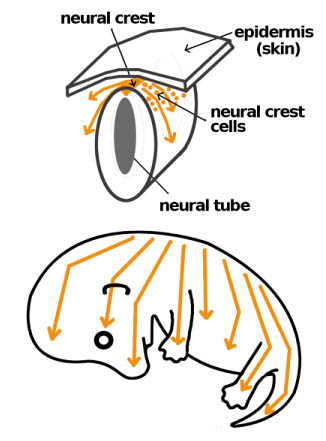
To understand how Miniature Schnauzers and all dog breeds arrive at the various colours and patterns they exhibit, we'll begin at the beginning. In the bottom layer of the skin's epidermis are cells called melanocytes. Melanocytes are derived during fetal development from the neural crest. In addition to the pigment producing melanocytes, other cell types generated from the neural crest include those making up the nervous sytem, adrenal gland, and many of the skeletal and connective tisues in the head. What type of cell they become depends on what part of the body they migrate to. Melanocytes produce melanin, the pigment found in the hair, skin, nose and pads. In addition to the creation of colourful pigment, melanin is a "photoprotectant", absorbing harmful UV radiation. Melanin occurs in two forms:
|
|
Variations in pigmentation relate to the number, size and distribution of melanosomes. Melanosomes are a specialized structure within the melanocytes that inject tiny pigment granules into the hairshaft as it is formed in the follicle.
For the purposes of simplication, I'll refer to as eumelanin as "black" and pheomelanin as "tan".

The broad rainbow of colours we see in our purebred dogs are the result of differing genetic combinations that alter the intensity and patterning of these two pigments. Thus, black pigment can be altered to brown, tan pigment can range from red to nearly white, and so forth.
A brief review of basic canine genetics.
 (Human chromosomes)
(Human chromosomes) |
Genes are arranged onto chromosomes. Chromosomes function in pairs, one inherited from the dam and the other from the sire. There are 39 pairs of chromosomes (for a total of 78) in the canine genome. Each gene occupies a specific location, or locus on its particular chromosome and codes for a specific cell protein or function, known as a trait. |
|
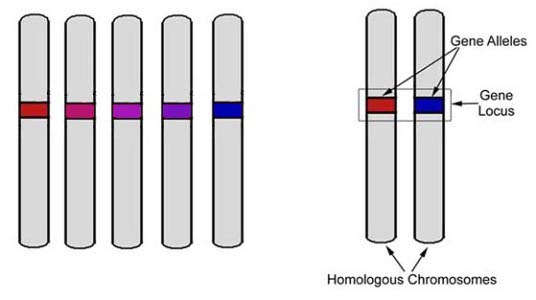
It's important to understand that while an individual dog can carry only two copies of a gene of any particular locus, within the gene pool of that breed may be several mutations of that gene (called alleles) that are capable of expressing in a variety of ways according to how they're combined. |
|
|
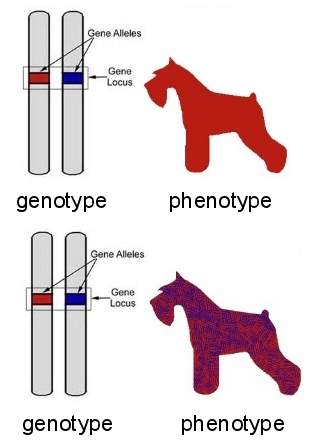
The genetic makeup of an individual is called the genotype. The observable traits that are expressed in that individual is the phenotype. When a copy of the gene inherited from father and mother are identical, they are called homozygous. When the gene copies inherited from the father and mother are different (different alleles), they are heterozygous. When only one allele in a heterozygous pair is expressed in the phenotype, we consider it to be dominant over its recessive partner. In other cases, the effect is only partial. Co-dominance, incomplete dominance, or intermediate expression can result in a shared or delayed expression of observable traits. One well-known case of this phenomenon is illustrated in the coat colour of Shorthorn cattle. When a calf born of a red bull and white cow receives a gene for each colour, the result will be "roan", a mixture of red and white hairs. |
|
In other cases, incomplete dominance can result in a delay in the expression of a trait, a phenomenon that we see in a number of disease defects that lay dormant until middle age. (Note that in some cases, a gene we describe as "dominant" can be actually be non-functional, paired with a normal gene that is incapable of "sustaining cell function on its own.)
|
|
Genes can even express in a variety of ways when arranged in identical combinations! For example, while all merle shelties are Mm at that locus, (indicating inheritance of two different alleles, or heterozygous) the patterns can vary greatly from dog to dog. |
 Both of these shelties are "Mm" at the merle locus
Both of these shelties are "Mm" at the merle locus{kind=link}
Even in cases of embryonic clones, white markings can vary. This is because the migration of melanocytes is essentially random, and may even be influenced by factors such as fetal position, nutrition of the dam, etc. |
 clones clones |

|
Finally, in rare cases coat colour can be affected by somatic mutations. Unlike mutations in the reproductive sperm and egg cells, somatic mutations occur after fertilization. For example, should a skin cell in a yellow lab embryo mutate from "ee" yellow to "Ee" black early in fetal development, the descendants of that cell will also be black, and express in the phenotype as random black patches. In general, dogs with mismarks caused by somatic mutations can't pass them on to their offspring. |
It is also important to know that many of the colours and patterns we see in dogs are the result of chemical processes that have profound effects on canine physiology, some of which are just being discovered. The most well known example is the extreme white piebald patterning seen in white bull terriers, dalmatians and a number of other breeds. The processes that create this extreme lack of pigment sometimes also cause deafness. Collie breeders know that doubling up on the gene for merle can result in puppies who are born without eyes. Many breeds with dilution colour genes suffer associated hair loss.
An aside to those of us concerned with animal rights encroachments on our liberties. It is imperative that breeders familiarize themselves with these issues so that we can mount reasoned arguments against accusations that breeding merles, etc. is inhumane. It is not good enough to say that colour genes dog breeders select for are not potentially harmful, when the truth is that some are.
Next...